In prehistory, the kinship systems that may have existed within communities can only be studied thanks to the archaeological and biological remains discovered during excavations. The study of isotopes, sex and age of buried individuals can provide information on the demographic structure of the population studied when it constitutes a funerary community. Genetics also plays a role in these studies, making it possible to determine family relationships, for example.
This is the conclusion of a study recently published in the journal Nature, which analyzed the genomes of over 100 individuals from the Gurgy “les Noisats” burial site (France). The site has been dated to between 4850 and 4500 BC.
The study highlighted several aspects of the community’s social organization. Thus, there are 2 main family groups: Group A, comprising 64 individuals, all related to each other and spanning 7 generations, and Group B, comprising 12 individuals, also all related to each other and spanning 5 generations.
The different generations are mainly linked by male ancestry. In fact, all descendants are linked by the paternal line of the first generation. Furthermore, with the exception of 2 individuals, no adult mothers have relatives or ancestors buried in this community. This, coupled with the fact that very few female individuals are descendants of the two main groups, suggests that this community was patrilineal, with an exogenous origin of the women in the group.
As far as the organization of burials is concerned, it has been shown that family ties influenced their layout. For example, burials between a father and son are physically much closer than those of other family members. The same applies to relationships other than father-son. The4th generation siblings were all buried close together. In another example, an adult mother’s son was buried above her. In this way, the spatial organization of burials forms organized groupings according to the more or less close kinship links between individuals.
Finally, the high number of adult siblings suggests a relatively low mortality rate among children.
This burial site was only occupied for a few decades.
The theory of evolution, which postulates that living beings evolve over time, emerged in the second half of the 19th century.
Until the 18th century, the predominant theory was that of fixism, which held that living beings had been created by the hand of God and were therefore immutable. For example, a name often associated with fixism is that of Carl von Linné (1707-1778). For him, all existing species were created during a single divine creation. Linnaeus is best known for his Systema Naturae. The most famous edition is that of 1758, in which he created our species, Homo sapiens, and classified it among the primates. However, for him, this classification of species has only one objective: to make intelligible the divine plan of Creation, of which human beings are the culmination.
Georges Cuvier (1769-1832) developed the theory of catastrophism. According to him, there is no evolution of living beings, but rather a succession of catastrophes and mass extinctions. These “catastrophes” would be followed by new divine creations. This explains the presence of fossils of now-extinct animal species found in ancient geological layers.
The first theory of evolution, in the general sense of changes in species over time, was formulated in the 18th century by Jean-Baptiste de Lamarck (1744-1829). This theory is known as transformism. According to Lamarck, species change over time. There is thus continuity between extinct fossil species and present-day species. Physical factors are thought to be at the origin of this modification of living beings, and the acquired traits are then passed on to offspring. Although this theory is erroneous, it is nevertheless the first theory in the direction of species evolution, representing an important step towards evolutionism.
Transformism according to Lamarck – the example of the giraffe’s neck
To illustrate his theory, Lamarck used the now-famous example of the evolution of giraffe neck size. In his work, Philosophie Zoologique, published in 1809, he explains:
“In terms of habits, it is curious to observe the product in the particular shape and size of the giraffe(camelo pardalis).): we know that this animal, the largest mammal, inhabits the interior of Africa, and that it lives in places where the earth, almost always arid and without grass, obliges it to graze the foliage of the trees, and to continually strive to reach them. As a result of this habit, which has been sustained for a long time in all individuals of its race, its front legs have become longer than its hind legs, and its neck has lengthened so much that the giraffe, without standing on its hind legs, raises its head to a height of six meters.
Philosophie Zoologie, book I, chapter VII.
Here, the environment determines the evolution of the giraffe’s neck size. It’s the function that shapes the organ. Because the giraffe needs to reach high up on leaves, its neck has lengthened, which over time has led to a change in the species. This acquired trait is then passed on to descendants.
Charles Darwin, Alfred Wallace & natural selection
The theory of evolution by natural selection was developed independently by Charles Darwin (1809-1882) and Alfred Russel Wallace (1823-1913) in the second half of the 19th century. Darwin developed this theory through his explorations aboard the Beagle, on which he spent 5 years (1831-1836). However, it wasn’t until 1859, 23 years after his round-the-world voyage, that Darwin published his famous book On the Origin of Species by Natural Selection or the Preservation of Favored Races in the Struggle for Life.
For the record, Darwin was prompted to publish his manuscript when he discovered that Wallace was about to publish a theory similar to his own. Finally, Darwin and Wallace’s work was presented on the same day to the London Linnaean Society in 1858. Nevertheless, it’s Darwin’s name that has gone down in history.
What is the natural selection theory of evolution?
Following his observations in the field, Darwin developed the following reasoning. First, he notes that there is variability among individuals of the same species. What’s more, these variations are heritable and selectable. Darwin arrives at this conclusion by observing the breeding practices and selection for reproduction that are applied to some livestock. In this way, he notes that selected traits are passed on to offspring. The question then arises: does this selection also take place in the wild, or only in a breeding context where breeders play the role of selection agents? Darwin identified that this selection agent is present in nature and corresponds to the environment in which individuals live.
There may be several variants for a single trait, and each variant is carried by several individuals. If environmental conditions favor one or more of these variants, individuals carrying them tend to leave more offspring than others. This differential reproductive success is what we call natural selection. On a population scale, this natural selection leads to transformations within species. The characteristics of advantaged individuals eventually become dominant within a population, as these individuals have a higher reproductive rate. This leads to a change in the population over time.
We can summarize the theory of evolution by natural selection as follows: individuals carrying an advantageous mutation in an environment will leave more descendants than other individuals not carrying this mutation. If the environmental conditions favorable to this mutation persist over time, the frequency of the favored variant in a given population will eventually reach 100%. As a result, the population, if not the species, will have changed. Subsequently, new mutations will occur for the same trait, leading to a new modification of the species if the environment is favorable. It is important to note that these mutations are the result of chance. The survival of a species is thus directly linked to its ability to have many variations for a single trait.
Work in genetics in the first half of the 20th century led to the discovery that these variations and mutations are genetic in origin, and that evolution therefore results from a change in allele frequency within a population. In short, mutations, which occur by chance, are the fuel of evolution, while natural selection is its driving force. Natural selection is at the root of a population’s adaptation to an environment.
Darwin’s example of the giraffe’s neck
Darwin uses Lamarck’s example of the giraffe’s neck:
“The high stature and elongation of the neck, forelimbs, head and tongue are conditions in the giraffe that adapt its entire frame admirably to the habit of grazing on the high branches of trees. […] in that individuals with one or more parts more elongated than usual have generally been the only ones to survive. Their cross-breeding has produced descendants either inheriting the same bodily characteristics, or a tendency to vary in the same way, while individuals less favored in the same respects will have been more likely to perish.”
The Origin of Species, 6th edition, 1872.
The main difference between Lamarck and Darwin lies in the mechanism underlying the transformation of species. According to Lamarck, it is environmental conditions that induce the modification of a character. In the example of the giraffe, the need to stretch the neck to reach the leaves directly lengthens the neck. This acquired physical trait is then passed on to offspring.
For Darwin, on the other hand, the driving force behind the transformation of species is natural selection. Giraffes with a genetic variation resulting in a longer neck have an advantage in their environment, as they can access food more easily. This advantage increases their chances of survival and, consequently, their ability to reproduce. As giraffes carrying this mutation have more offspring, its frequency increases in the population, leading to a change in the population.
Figure 1: Diagrams showing Lamarck’s and Darwin’s theories of evolution.
The theory of evolution by natural selection at a glance
Here are a few points to remember if you want to fully understand the theory of evolution by natural selection:
Biological evolution applies not to an individual but to a population. Natural selection has an effect on individuals, but it’s a population that evolves and changes over time.
Evolution has no purpose; it is the result of the randomness of genetic mutations. There is no intention or will behind evolution.
Our species, Homo sapiens, is no more evolved than any other animal species. Homo sapiens is an animal, a mammal, a primate and an ape like any other (if you want to learn more about them, click here). We are not the end product of evolution, we are the result of chance.
The evolution isn’t over yet! All species are always evolving, including us! In reality, a species that doesn’t evolve is doomed to extinction. Consequently, the term “living fossils” is biologically meaningless.
Similarly, there are no evolutionary stages or steps in evolution, it’s a continuous process.
We hope you enjoyed this article! Feel free to ask us questions and give us feedback on the blog. You can also contact us by e-mail. You can also follow us on Instagram, Facebook, TikTok, Twitter, LinkedIn and YouTube!
See you soon,
The Prehistory Travel team.
Bibliography :
[1] Boyer Charles, “Le cou de la girafe : Lamarck, et puis Darwin”, L’Enseignement philosophique, 2011/2 (61e Année), pp. 48-54. DOI : 10.3917/eph.612.0048. URL: https://www.cairn.info/revue-l-enseignement-philosophique-2011-2-page-48.htm.
[2] Darwin Charles, On the Origin of Species by Natural Selection or the Preservation of Favored Races in the Struggle for Life, ed. orginale 1859, 2022, éditions Flammarion.
[3] Lamarck Jean-Baptiste, Philosophie zoologique ou Exposition des considérations sur l’histoire naturelle des animaux, tome I, ed. original 1809, 2017, Hachette Livre BNF.
How do you determine whether or not a fossil belongs to the human lineage?
It’s important to distinguish between two things: Hominins and the human line. Hominins include representatives of the Homo genus and all fossils closer to us than chimpanzees. The human lineage refers to the ancestors of our species, Homo sapiens. So, among the Hominins, paleoanthropologists are looking for those that could be at the origin of the human lineage. To do this, they defined criteria such as mode of locomotion.
Indeed, although some anatomical criteria exist, it is mainly bipedal characteristics that are used to determine whether or not a fossil belongs to the human lineage. Why bipedalism? Quite simply because it’s the preferred mode of locomotion for our species! We must be careful, however, as Homo sapiens is not the only species capable of bipedalism. In fact, the ability to move in a bipedal manner, i.e. on both lower limbs, appeared well before the emergence of the human lineage around 7 million years ago (later abbreviated to Ma). Indeed, this type of locomotion appeared as early as 250-200 Ma. It is found, for example, in certain dinosaurs. Even today, other animals such as birds and kangaroos move around the ground in a bipedal fashion. Among primates, the order to which Homo sapiens belongs, many species are capable of bipedalism. In fact, our species is the only primate to practice a single mode of locomotion for which our bodies are hyper-specialized. Furthermore, scientists agree to no longer speak of a single biped, but of bipeds, to emphasize that different bipedal locomotions have existed within the Hominins. To find out more about the skeletal adaptations required for bipedalism, click here.
Exclusive bipedalism therefore seems to be a feature of our lineage. This is probably why studies on the bipedalism of early Hominins are at the heart of the debate. What about our three candidates?
Sahelanthropus tchadensis, one of the oldest candidates for the origin of the human lineage
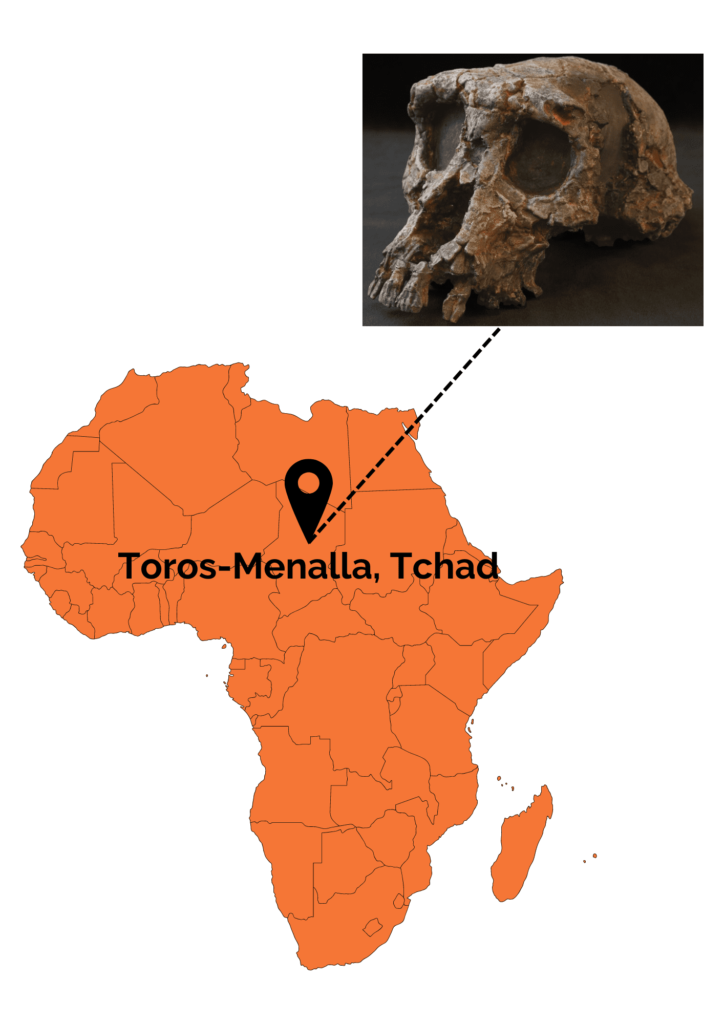
This species was discovered between 2001 and 2002 by a Franco-Chadian team at Toros-Menalla in Chad (Africa). Several cranial remains (3 mandibles and several isolated teeth), belonging to at least 3 different individuals, as well as an almost complete skull were unearthed. What’s more, this skull is nicknamed Toumaï! A few postcranial remains (a left femoral shaft, two right ulnas and a left ulna) were also discovered. The only problem with the latter remains…they were not discovered in direct connection with the cranio-dental remains. Consequently, it is impossible to state with certainty that they belong to S. tchadensis! Nevertheless, the discoverers of the fossils feel that the most parsimonious hypothesis is to consider that these postcranial remains belong to the only species present on the site, S. tchadensis.
Figure 1: Location of the Toros-Menalla site and photograph of the Sahelanthropus tchadensis skull. Frank Guy.
The fossil remains of S. tchadensis have been dated to between 7 and 6 Ma using two methods. The first is biochronology, i.e. a chronological estimate based on the degree of evolution of fossil faunal remains found within the same stratigraphic zone. The second method used is cosmogenic nuclide dating (Aluminum 26Al/Beryllium10Be). According to the fauna associated with the fossil remains and the palynological studies carried out in the region, S. tchadensis lived in an environment combining forests, savannahs, grasslands and bodies of water. This region is now desert.
Several anatomical features bring us closer together S. tchadensis of later Hominins, such as small canines, absence of diastema (space between incisors and canines due to the large size of the latter) and a more anteriorly positioned foramen magnum than in non-human great apes. These characteristics distinguish S. tchadensis from gorilla ancestors, a hypothesis put forward at the time the fossils were discovered. Nevertheless, the cranial features must be interpreted with caution, as the skull is deformed by the weight of the sediment. As a result, certain characters have been deduced from virtual reconstructions. Other features, such as a small cranial capacity of around 350 cm3, bring S. tchadensis closer to non-human primates.
Figure 2: Morphological features of the S. tchadensis skull. Institut Paléonthologie Humaine
Toumaï is far from being unanimously accepted by the scientific community. For its discoverers, the anterior position of the foramen magnum indicates bipedal locomotion. For many other scientists, however, this alone is not enough, especially as the base of the skull, where the foramen magnum is located, is severely damaged. What’s more, the few postcranial remains found, notably the femurs, do not seem to reveal any clear adaptations to bipedalism. The debate continues with a focus on dental residuals. Apart from bipedalism, the size of the canines (reduced in H. sapiens) is also a criterion used to assign a fossil to the Hominins. S. tchadensis has a priori small canines, but only if we consider that the skull found belongs to a male. This deduction is based solely on the size and width of the supraorbital torus. However, the link between supraorbital torus size and gender is far from unanimous, as it is considered an unreliable trait. For some researchers, the most prudent hypothesis would therefore be to consider that the skull belongs to a female. The small size of the canines would therefore not be the result of any link with Hominins, but simply linked to a phenomenon of sexual dimorphism.
The saga continues with a new article published in 2022 in Nature where a femur, discovered more than 20 years ago, has just been officially described and whose study shows that a dozen characters demonstrate the bipedal ability of S. tchadensis although he was certainly still climbing trees. Finally, in 2023, Marc R. Meyer et al. published a study that raised the hypothesis of knuckle-walking locomotion! This would exclude S. tchadensis from the Hominins.
As you can see, although still considered a potential ancestor of the human lineage, the position of S. tchadensis is hotly debated.
Orrorin tugenensis
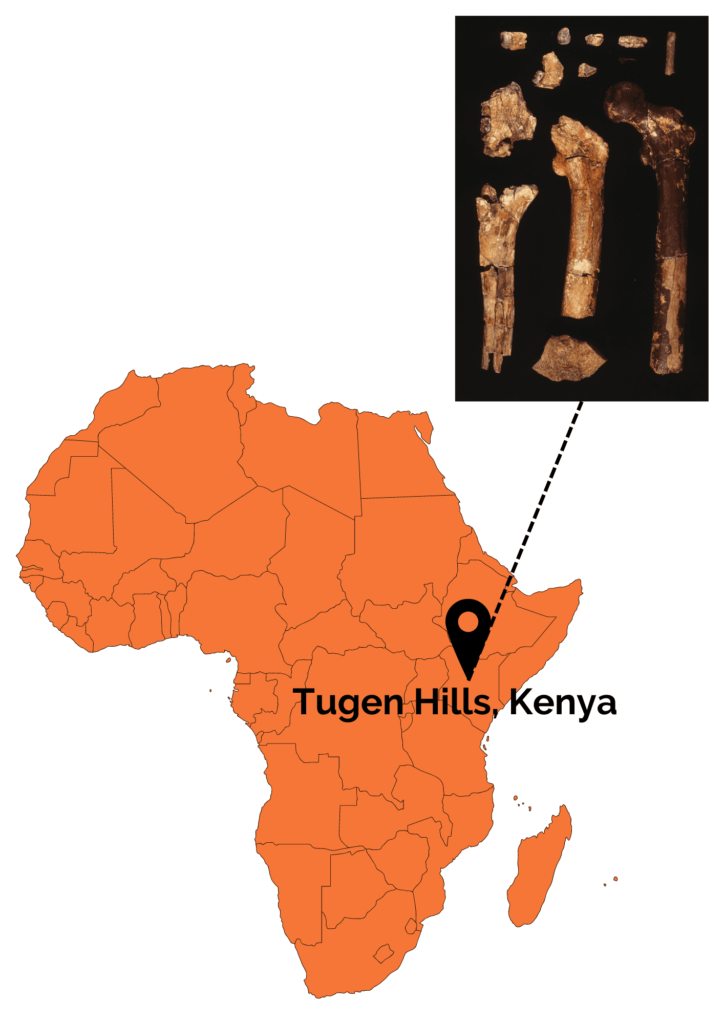
The species Orrorin tugenensis was created in 2000 following the discovery of a dozen dental and postcranial remains on Tugens Hill in Kenya by Martin Pickford and Brigitte Senut. These remains are dated between 6.1 and 5.7 Ma.
Figure 3: Location of the Tugens Hill site and photographs of the fossil remains associated with Orrorin tugenensis. Martin Pickford.
Several morphological features bring O. tugenensis closer to the more recent Hominins. In terms of dentition, the molars found are square-shaped and small compared to other non-human primates. In addition, the canines are reduced in size and there is an absence of diastema. Unlike chimpanzees, Orrorin’s upper molars show different degrees of wear, suggesting differential molar growth. In fact, the first molar, which is the most worn, would have come out before the second molar and finally the third molar, which is hardly worn at all. This differentiated growth of molars is found not only in H. sapiens, but also in Australopithecines.
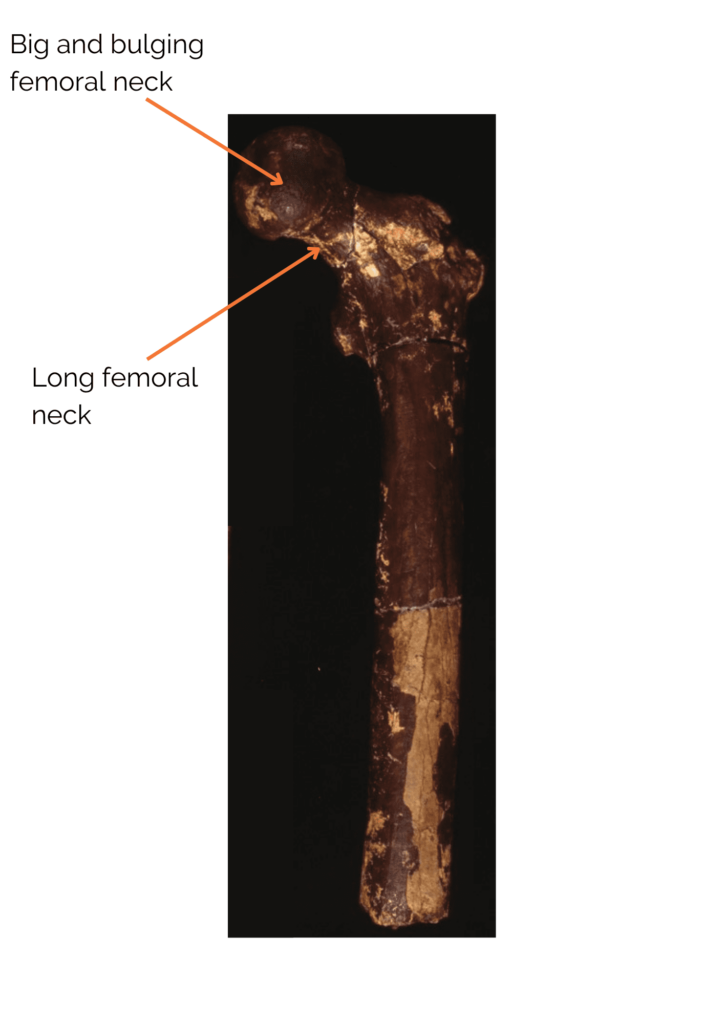
Postcranial remains show several signs of regular bipedal locomotion. These include a long femoral neck with a large, globular head, similar to that of Homo sapiens. In addition, the cortical bone at the femoral neck is very thick. This is a good indication of bipedalism, as it is interpreted in bipeds as reinforcement to support the weight of the trunk without risk of breakage. Nevertheless, although Orrorin was probably an occasional or regular biped, his bipedalism was probably not like ours. It’s also worth noting that the bones of the upper limbs, notably the curved phalanx of the hand, show adaptations to tree climbing. Orrorin was therefore capable of both arboricolia and bipedalism! In terms of its environment, studies have shown that Orrorin evolves in a dry temperate forest environment with the presence of wetlands. This has been much debated within the scientific community, as it contradicts the theory of the emergence of bipedalism through body straightening due to an open environment. Studies of its teeth suggest that it was a frugivore or omnivore.
Figure 4: Femur ofOrrorin tugenensis. Martin Pickford.
The Ardipithecus genus
The latest candidate for the title of oldest Hominin is the genus Ardipithecus. Two species belong to this genus:
Ardipithecus kadabba, dated to between 5.77 and 5.2 Ma, whose fossils have been found in various localities in the middle Awash valley in Ethiopia.
Ardipithecus ramidus, also discovered in Ethiopia’s Middle Awash Valley and dated at around 4.4 Ma
Figure 5: Location of the Middle Awash Valley in Africa.
A. kadabba has many primitive features, such as large canines implying the presence of a diastema, and a particularly strong elbow joint indicating brachiation locomotion. Similarly, the phalanges of the hands and feet are long and curved, indicating a capacity for arboricolia. Nevertheless, there are a few features that bring this species closer to the human lineage. This is the case, for example, of the articular surface of a proximal foot phalanx, which has a dorsal inclination. This trait is considered unique to bipeds, linking A. kadabba to more recent Hominins. A. kadabba is sometimes seen as the ancestor of the second known species belonging to the genus Ardipithecus: Ardipithecus ramidus.
Like its potential ancestor, Ardipithecus ramidus also displays features that indicate both arboreal and bipedal locomotion.
Although the place of these 3 genera within the Hominins remains debated, their importance in understanding the origin of the human lineage is certain! Indeed, these specimens are from a period when our ancestors diverged from the ancestors of chimpanzees, our closest cousins, and thus illustrate the beginning of human history.
[1] Michel Brunet, “Sahelanthropus tchadensis dit “Toumaï” : le plus ancien membre connu de notre tribu”, Bulletin de l’Académie Nationale de Médecine, 204 (2020)
[2] Michel Brunet et al, “Toumaï, Miocène supérieur du Tchad, le nouveau doyen du rameau humain”, Comptes Rendus Palevol, 3 (2004).
[3] G. Daver et al, “Postcranial evidence of late Miocene hominin bipedalism in Chad”, Nature, 609 (2022), https://doi.org/10.1038/s41586-022-04901-z
[4] Martin R. Meyer et al, “Knuckle-walking in Sahelanthropus? Locomotor inferences from the ulnae of fossil hominins and other hominoids”, Journal of Human Evolution, 179 (2023), 103355
[5] M. Pickford et al, “Bipedalism in Orrorin tugenensis revealed by its femora”, Compte rendus Palevol, 1 (2002)
[6] Y. Sawada, “The age of Orrorin tugenensis, an early hominid from the Tugen Hills, Kenya”, Comptes rendus Palevol, 1 (2002)
[7] B. Senut, ” Orrorin tugenensis et les origines de l’homme: une synthèse”, Bulletin de l’Académie Nationale de Médecine, 204 (2020)
[8] Peter A. Stamos et al, “Hominin locomotion and evolution in the Late Miocene to Late Pliocene”, Journal of Human Evolution, 178 (2023), 103332
[9] Tim D. White et al, “Ardipithecus ramidus and the Paleobiology of early hominids”, Science, 326 (2009)
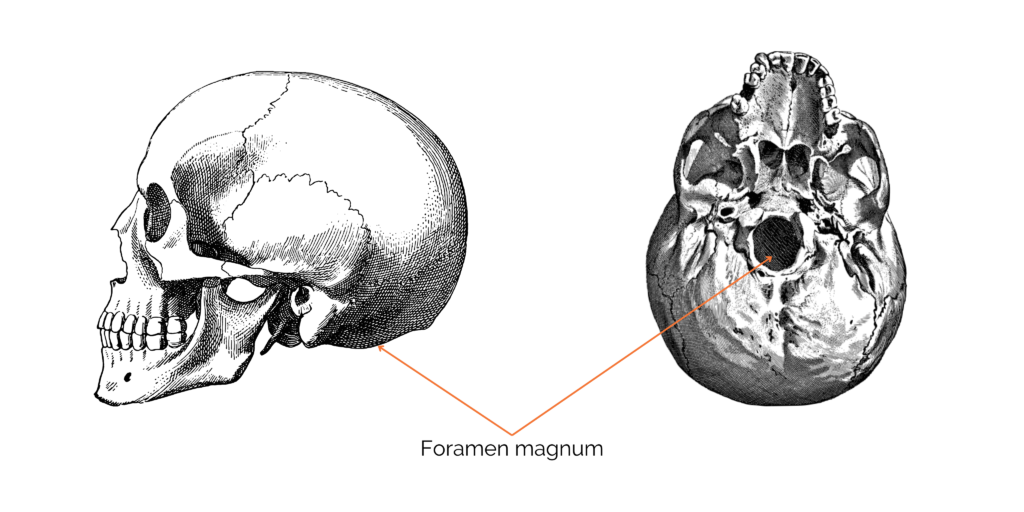
In Homo sapiens, the head rests on top of the spinal column via the foramen magnum. The foramen magnum, positioned under the skull, allows the spinal column to be attached and the medulla oblongata to pass through. The medulla oblongata forms the lower part of the brain stem. Its position under the skull is very important, as it gives indications of the general position of the body and therefore of the mode of locomotion. In our species, the foramen magnum is located in the center of the skull and is oriented upwards and forwards, enabling us to keep our heads upright during bipedalism. In other primates, the foramen magnum is in a more posterior position, pointing downwards.
Figure 1: Position of the foramen magnum in Homo sapiens.
Figure 2: Comparison between the skull of a Homo sapiens (left) and a chimpanzee, Pan troglodytes (right).
Skeletal adaptations in the spine
The spine of Homo sapiens has a quadruple curvature: sacral kyphosis, cervical lordosis, thoracic kyphosis and lumbar lordosis. In non-human primates, the spine has a curvature. In the chimpanzee(Pan troglodytes), for example, cervical lordosis is less pronounced, and there is only a single curvature for the rest of the spine. This specificity in today’s human beings allows for greater muscular efficiency in maintaining the body upright. This also provides greater resistance during bipedal gait.
Figure 3: Spine in Homo sapiens (A) and in a non-human primate (B).
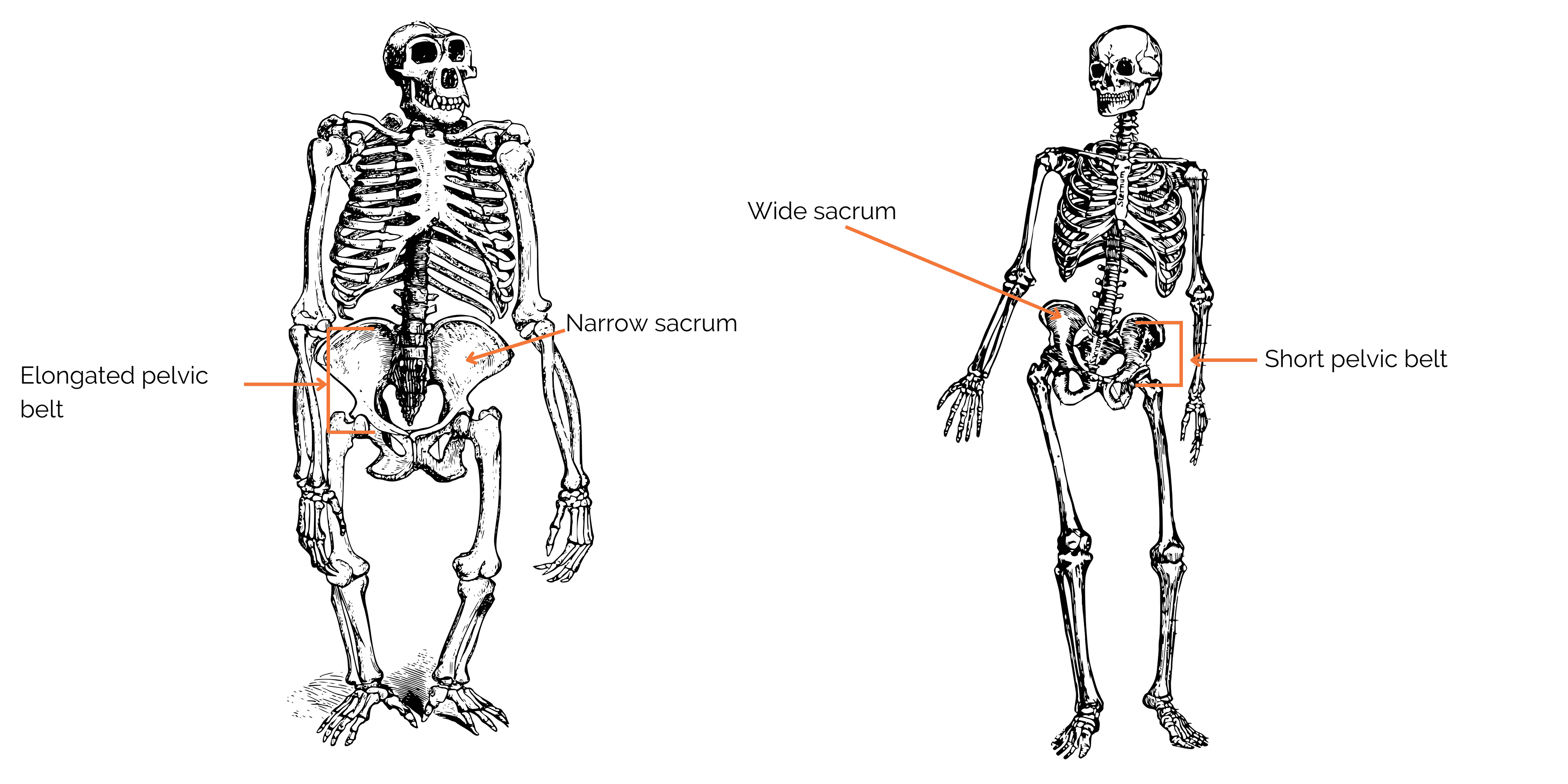
Skeletal adaptations in the pelvis
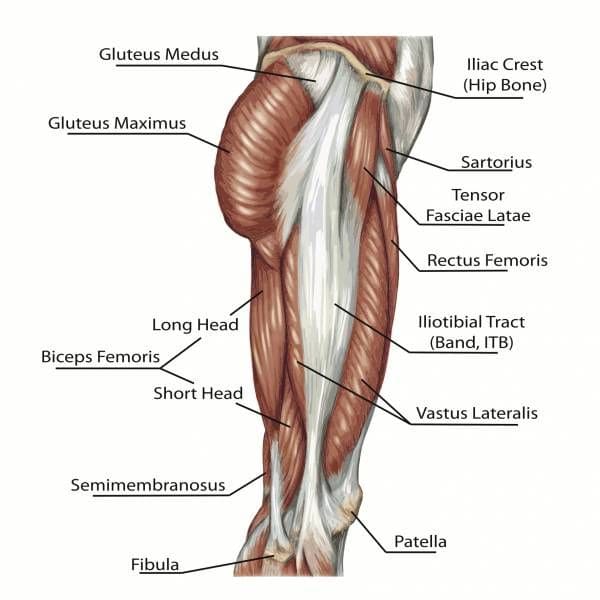
In non-human primates, the pelvis is very elongated. In human beings, the acquisition of bipedalism has been accompanied by a widening and shortening of the pelvis (known as compression). This remodeling offers numerous biomechanical advantages, such as visceral support. This enables optimal positioning of the gluteus medius muscle (formerly gluteus medius), which becomes a thigh abductor rather than a rotator as in other primates. This limits trunk oscillations in the frontal plane (to the sides), compared with the bipedal, shuffling gait of chimpanzees. On the other hand, in the sagittal plane (direction of travel), our center of mass oscillates, allowing us to recuperate our energy and thus have a very efficient bipedal gait!
The ischial tuberosities have also migrated posteriorly in humans, providing the hamstrings with optimum leverage for the erect bipedal posture. What’s more, the lumbar spine and pelvis serve as anchors for the trunk muscles, keeping the trunk optimally erect and requiring little muscular effort.
Figure 4: Comparison between the pelvis of a chimpanzee (left) and that of a Homo sapiens (right).
Figure 5 : Diagram of gluteal and upper thigh muscles in modern humans.
Skeletal adaptations in the lower limbs
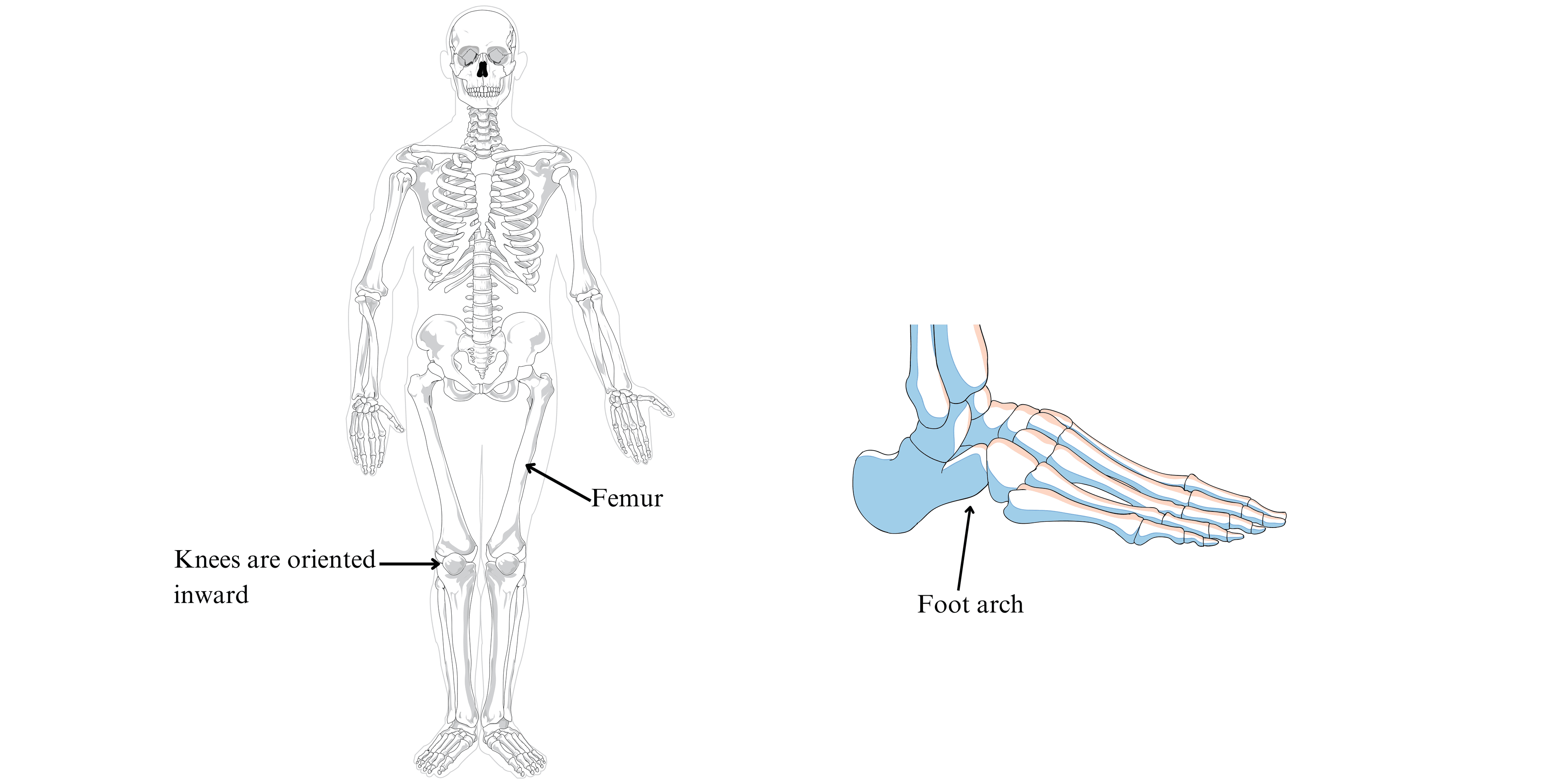
With bipedalism, the lower limbs must carry the entire weight of the body. These limbs are longer in H. sapiens than in other primates. In addition, the femurs are oblique and the knees inward, bringing them closer to the body’s line of gravity. Knee and ankle joints are reinforced for greater stability. The joints of the lower limbs function mainly in the parasagittal plane (direction of gait).
The foot is hollowed out by two arches, one transverse and the other longitudinal, stabilizing the foot on the ground and transferring weight from the heel to the forefoot. The big toe is aligned with the other toes, which, among primates, is unique to our species! The toes are short and the heel is massive.
Figure 6:Homo sapiens skeleton and diagram of the foot with longitudinal arch.
Here are a few examples of the musculoskeletal adaptations needed to enable us to walk upright efficiently for long periods. Bipedalism is at the heart of the debate on the emergence of the human lineage. Indeed, this is considered one of the main criteria for determining whether an extinct species whose fossil remains have been found belongs to the Hominins or not.
We hope you found this article interesting. Feel free to ask us questions and give us feedback on the blog. You can also contact us by email. Follow us on Instagram, Facebook, Twitter, TikTok,LinkedIn and YouTube to keep up with all the latest news!
We would like to thank François Druelle, a research fellow specializing in primatology and locomotion, for reviewing the first version of this article.
A 2-million-year-old protein sequence (later abbreviated to Ma) has just been successfully sequenced and analyzed! This is the conclusion of an article published on the bioRxiv server, which allows articles to be put online before the peer review process is completed on July 3, 2023. These protein sequences come from several teeth of individuals belonging to the Paranthropus robustus species. These teeth were found in a cave in South Africa. This is an exceptional discovery. In fact, studies on ancient DNA do not allow us to go back as far as 2 Ma, as DNA does not keep well over time and is particularly sensitive to heat. For example, the oldest sequenced hominin DNA is dated to around 430,000 years ago in Europe. We’re a long way from 2 Ma!
In recent years, the study of protein sequences has emerged as an alternative to the difficulties encountered in paleogenetics. Proteins are better preserved over time and directly reflect the genome, since they are made from our DNA sequence. In this study, the researchers succeeded in determining the sex of the Paranthropus individuals studied. In fact, they found traces of a protein called Y-amelogenin on 2 teeth. This protein is produced by a gene on the Y chromosome, indicating that these two individuals are male. In contrast, the other two teeth do not display this protein, but rather a protein present only on the X chromosome, suggesting that the teeth belong to females. Such a breakthrough in paleoproteomics looks promising. However, care must be taken, as this article is not “officially” published in a journal, as it has not yet passed the peer review stage, i.e. verification by other researchers of the quality of the study carried out.
An article was recently published in the journal PlosOne reporting the discovery of engravings made by Homo neanderthalensis at the La Roche-Cotard cave. This cave, located in the Touraine region, more precisely at Langeais in Indre-et-Loire, was discovered in 1846. Several excavations were carried out on this site in 1912, in the 1970s and in 2008. This study reveals that the cave was first occupied by carnivores, then by humans and finally by hyenas, before the entrance was blocked and the cave became inaccessible until its discovery in 1846.
The human beings who lived in this cave were Neanderthals. How do we know? Thanks to dating, scientists have been able to determine that the cave entrance was blocked by sediment around 57,000 years ago. Consequently, this cave was occupied over 57,000 years ago, and at that time our species, Homo sapiens, had not yet arrived in Europe, the only species present being Homo neanderthalensis. What’s more, the tools found at the site are typical of the Mousterian, a culture strictly associated with Neanderthal in Europe.
Several marks and traces have been identified on the cave walls. These have been classified according to their origin. This ranking was based on experiments and statistical studies. Some of the marks are carnivore claw marks, but some are man-made and correspond to digital tracings. Digital tracings are lines or geometric shapes created with the fingers on an initially soft surface. This type of tracery is part of the various forms of cave art and can be found at other sites in Europe and South Africa, for example. The authors conclude that these tracings are the result of a creative process within the cave. This discovery proves, if proof were needed, that Neanderthals were capable of artistic, and perhaps even symbolic, practices!
Wiseman LA. 2023 Three-dimensional volumetric muscle reconstruction of the Australopithecus afarensis pelvis and limb, with estimations of limb leverage. R. Soc. Open. Sci. 10:2303556.
Researchers have reconstructed the muscle mass ofAustralopithecus afarensis Lucy, dated at 3.2 million years. This one, discovered in 1974 at the Hadar site in Ethiopia, provided us with a very complete skeleton, with almost 40% of the bones found. The various studies carried out on Lucy’s skeleton have shown that she was certainly capable of bipedalism, while continuing to climb trees to get around. However, these studies were carried out on bones, not muscles. Although researchers have been trying for some years to reconstruct the muscles of certain Hominins in order to deduce elements about their locomotion, this approach was limited by the complexity of the mathematical models to be set up.
In this new study, researchers try a new approach to reconstruct 36 muscles of the Australopithecus pelvis and lower limbs. To do this, they used the anatomy of Homo sapiens individuals to infer muscle positioning in Lucy. This new 3D model of Lucy’s muscles shows that she had much more massive and developed muscles than today’s Homo sapiens. What’s more, the study shows that, although certain muscles were positioned differently in Australopithecus than in our species, they enabled Lucy to stand in an erect position. However, his bipedalism differed from ours in the positioning of certain muscles. It was also capable of other modes of locomotion, such as arboricolgy.
The vast majority of primates live in tropical climates and forest environments. Some species also live in savannah-type environments or at higher altitudes. This is the case, for example, of the Barbary macaque, which lives in the Atlas mountains and on the promontories of Gibraltar. We can also mention the Roxellan Rhinopithecus, which lives on the Tibetan plateau, in a sometimes snowy environment. If you’d like to find out more about primates, we invite you to read our previous article. Thus, primates live both arboreal and terrestrial lives. This diversity of occupied environments leads to great diversity in primate postural-locomotor repertoires. Unlike most other mammals, primates can practice different modes of locomotion within the same repertoire. 32 posturo-locomotor modes have been identified, and primates are commonly grouped into ten major locomotor categories:
Vertical jumpers and climbers
Brachiators
General arboreal quadrupeds
Semi-terrestrial quadrupeds
Land quadrupeds
Quadrupedal knuckle-walking
Slow climbers
Claw climbers
Biped
Some American primates also have a prehensile tail, which they use as a fifth limb to suspend and stabilize themselves in trees.
Figure 1: Photograph of a red-handed tamarin, Saguinus midas, an American primate. Royalty-free photography.
These different locomotor modes are characterized by anatomical adaptations to the skeleton. However, we have to be careful when we talk about these adaptations. Indeed, primatologists now know that it’s not necessarily the time spent on the main mode of locomotion that best reflects skeletal adaptations. For example, the chimpanzee is defined as a knuckle-walker according to the broad categorization mentioned above. Nevertheless, most of the chimpanzee’s skeletal adaptations are not for this quadrupedal walking, but for suspension and vertical climbing, which it also practices. Categorizing posturo-locomotor modes helps us to define them. Nevertheless, it’s important to bear in mind that each primate species has its own way of doing things.
In the rest of this article, we’ll take a look at some of these modes of transport.
Vertical jumpers and climbers
Tarsiers, for example, use this mode of locomotion. The term “jumper” comes from their movement through the trees and on the ground in successive leaps. They can leap a distance that is up to forty times greater than their size! The term “vertical climbers” derives from the fact that tarsiers climb trees vertically, straightening their backs and spreading their limbs to cling to the trunk. The main adaptations to this type of locomotion are found in the postcranial region, and more specifically in the lower limbs, which are particularly elongated. In fact, they are 1.5 times the length of the trunk, providing support for propulsion during jumps.
Brachiation is defined as bimanual progression over several meters between two structures without using any other type of locomotion and without using the support of the tail or hind limbs. This type of locomotion is used by gibbons and siamangs, for example. Brachiator upper limbs are particularly long and strong. Their wrist and shoulder joints are also mobile. This type of locomotion enables them to move very quickly through the canopy – up to 56 km/h for gibbons!
Figure 3: Gibbon brachiating. Royalty-free video.
Quadrupedalism
As mentioned at the start of this article, there are three main modes of quadrupedalism: general arboreal quadrupeds, semi-terrestrial quadrupeds and terrestrial quadrupeds. They are all characterized by walking on all four limbs, supported by hands and feet. Quadrupeds can be digitigrades (supported by the toes of the hands and feet), semi-plantigrades, plantigrades (supported by the palms of the hands and feet) or a mixture of both! Baboons, for example, are digitigrade for their hands, but semi-plantigrade for their feet.
The distinction between these modes of quadrupedal locomotion depends on the environment in which they are practiced. Generalist arboreal quadrupeds, such as sakis and cercopithecines, move by walking or running on branches. Semi-terrestrial quadrupeds spend a variable proportion of their time in trees, and come down to the ground to feed. Macaques in general are a good example. Finally, terrestrial quadrupeds live mainly on the ground. Baboons are a case in point.
Figure 4: Photograph of a white-faced saki, Pithecia pithecia, moving in a quadrupedal fashion. Royalty-free photography.
Quadrupedal locomotion on the knuckles of the hands or knuckle-walking
Knuckle-walking is a form of locomotion that involves moving around on all four limbs, supported by the backs of the phalanges on the upper limbs. This type of movement is practiced by chimpanzees, bonobos and gorillas, for example. This requires adaptations to the bone structure of the wrist and metacarpals, which are more elongated and robust.
Figure 5: Photograph of a chimpanzee, Pan troglodytes. Royalty-free photography.
We hope you enjoyed this article! Feel free to ask us questions and give us feedback on the blog. You can also contact us by email. You can also follow us on Instagram, Facebook, Twitter, TikTok, LinkedIn and YouTube!
We would like to thank François Druelle, a research fellow specializing in primatology, for his review of the first version of this article.
See you soon,
The Prehistory Travel team.
Bibliography:
[1] Berillon G., Marchal F., “Les multiples bipédies des hominidés”, Pour la Science, vol. 330, 2005.
[2] Grimaud-Hervé et al, Ancestor stories. The great adventure of prehistoryed. Errances,5th edition, 2015.
[3] Kimura, “Habitual locomotor types and the shape of lower leg bones in primates, especially in hominoids”, Revue de primatology [en ligne], 12|2021.
[4] Michilsens et al, “Functional anatomy of the gibbon forelimb: adaptations to a brachiating lifestyle”, J Anat, 2009 Sep; 215(3): 335-354.
[5] Rasmussen et al, “Tarsier-like locomotor specializations in the Oligocene primate Afrotarsius”, Proc. Natl. Acad. Sci. USA, vol. 95, 1998
[6] Springer C., Andrews P., The complete world of Human evolution, ed. Thames & Hudson, 2011
[7] Tarrega-Saunders et al, “Knuckle-walking and behavioural flexibility in great apes”, Revue de primatology [en ligne], 12|2021
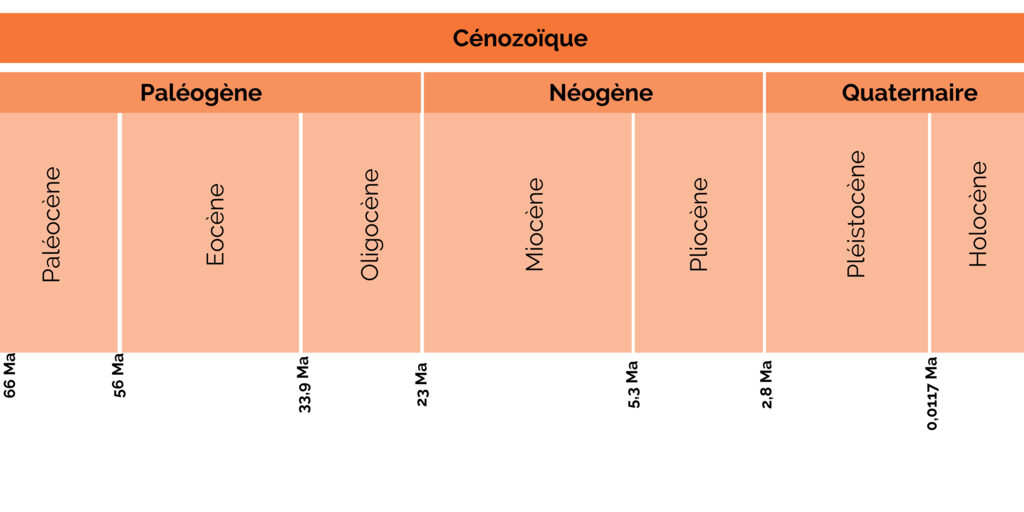
Once again, this article goes beyond the realm of prehistory. Although… the Cenozoic is the most recent geological era in which we are currently living, and it also includes prehistory! The Cenozoic is the era during which the Earth evolved into the one we know today.
Early Cenozoic
The Cenozoic era began with an extremely famous event: the extinction of the dinosaurs. Also known as the “K/T extinction” or “Cretaceous/Tertiary extinction”, this mass extinction took place 66 million years ago (later abbreviated to Ma). In fact, dinosaurs are not the only living creatures to disappear. Over 60% of the world’s flora and fauna are becoming extinct. Mammals appeared shortly before 200 Ma, during the era preceding the Cenozoic, and were small animals at the time. The massive disappearance of large aquatic and terrestrial animals such as dinosaurs has enabled mammals to flourish, freeing up new ecological niches. Why is this important? Keep in mind that without mammals, there are no primates, and without primates … noHomo sapiens, our species!
Figure 1: Geological division of the Cenozoic.
What happens during this period?
In general, the Cenozoic was marked by intense tectonic activity and intensified orogenesis, particularly in the Alps and Pyrenees. These gave rise to today’s mountain ranges. Continental drift has also led to their current position. Climatic instability sets in throughout the Cenozoic, resulting in the alternation of glacial and interglacial periods.
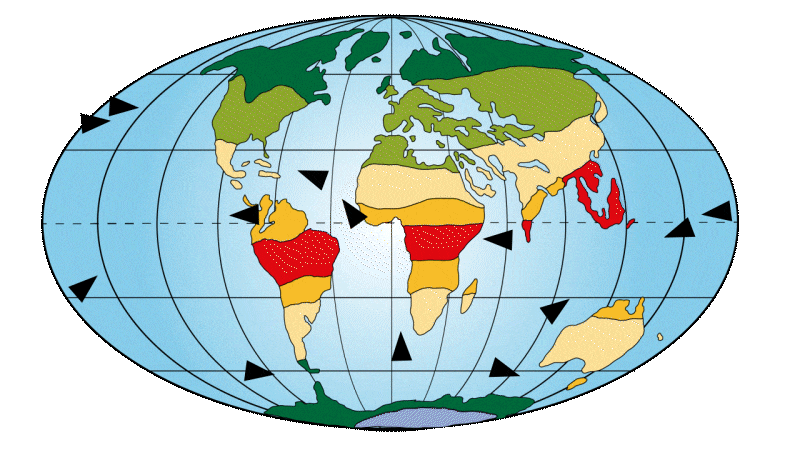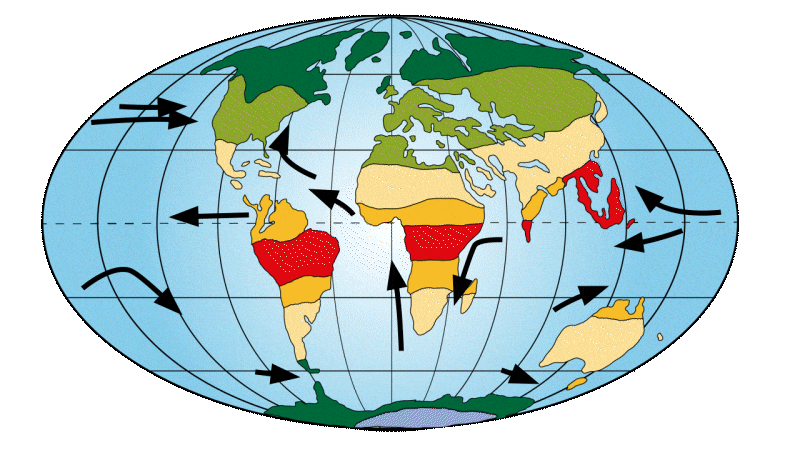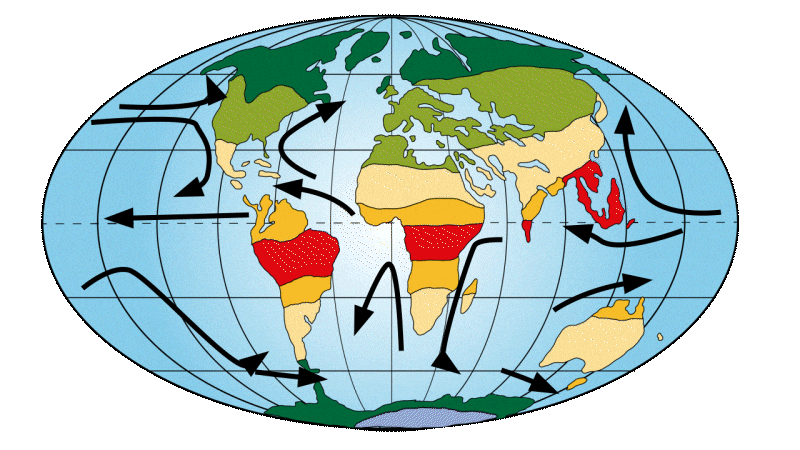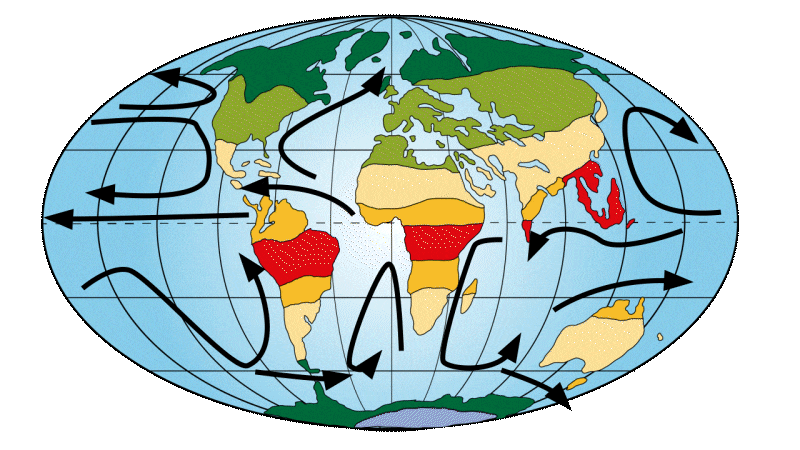
Around 65 million years ago
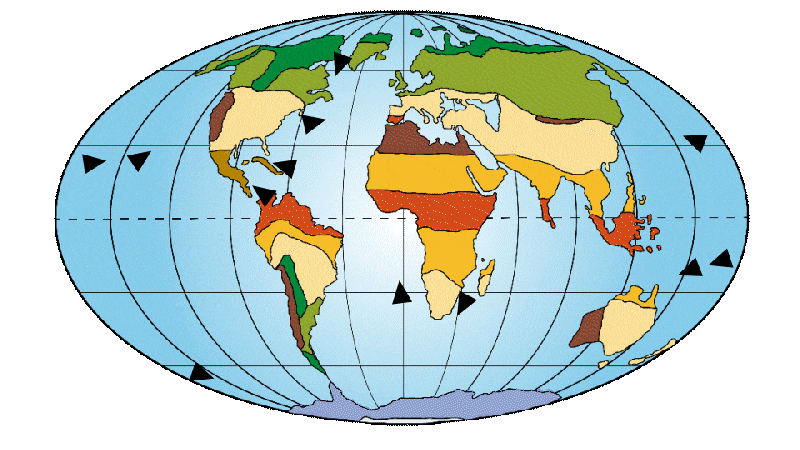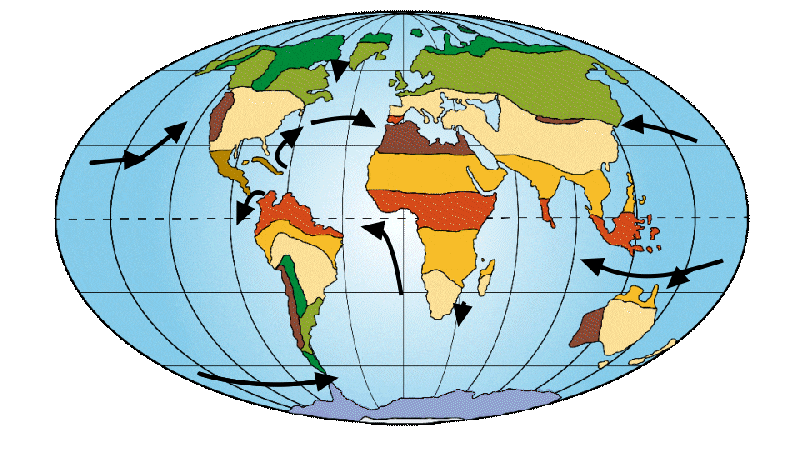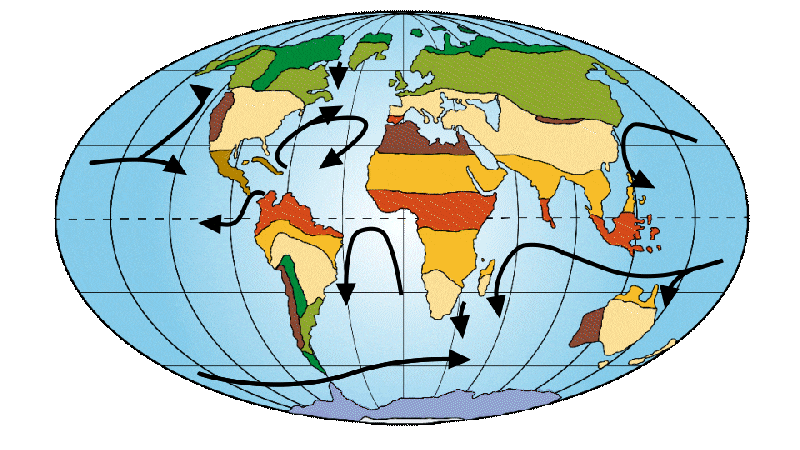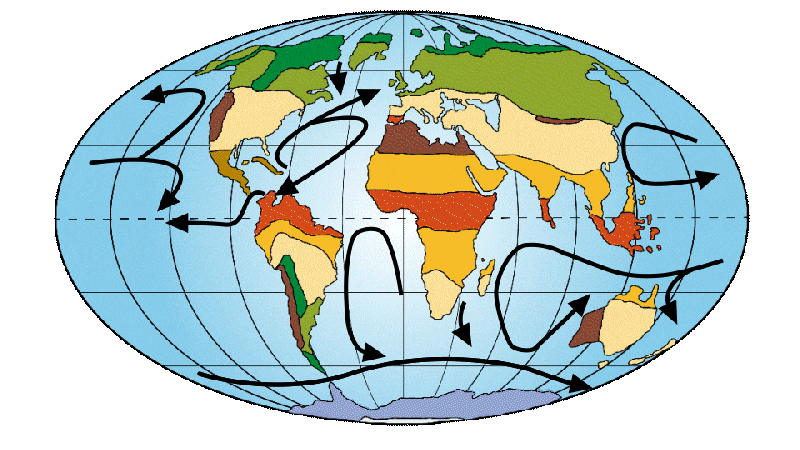
The continents are still relatively well centered on the equator and divided between the northern and southern hemispheres. This creates a large equatorial ocean current, homogenizing ocean temperatures towards the warm side. As a result, tropical and subtropical regions account for the majority of our planet’s population. Many land bridges exist at this time, helping mammals to disperse around the globe. One example is the area linking North America and the Eurasian continent, known as “Beringia”.
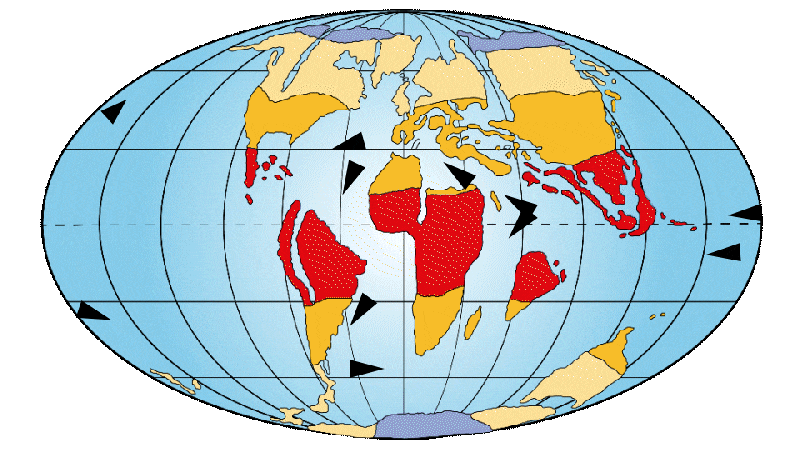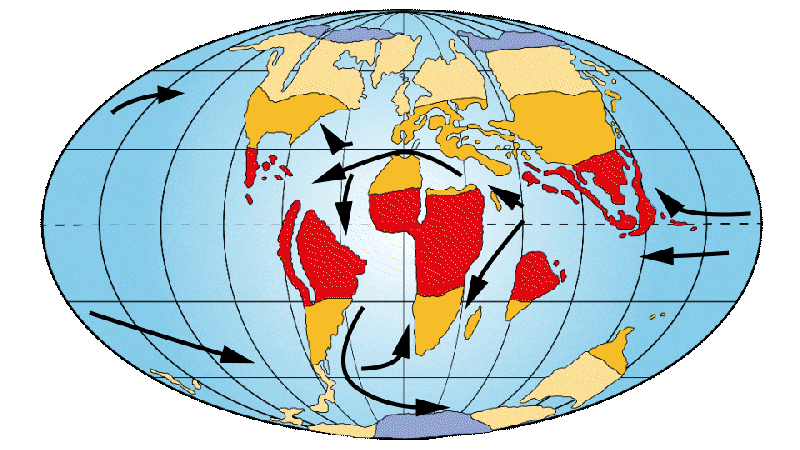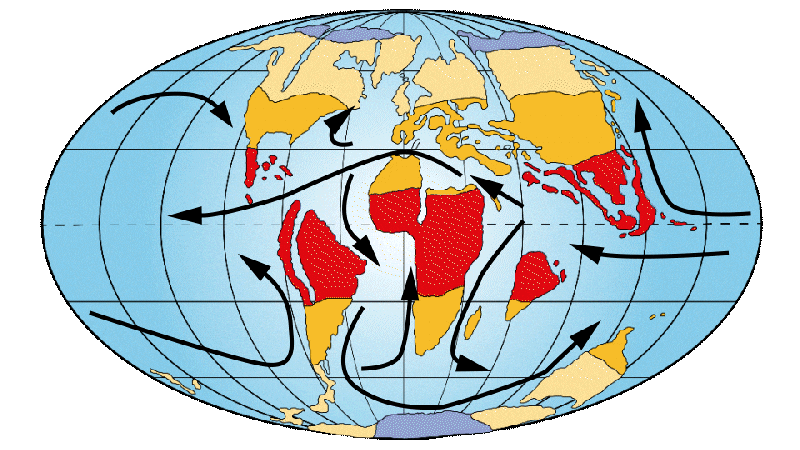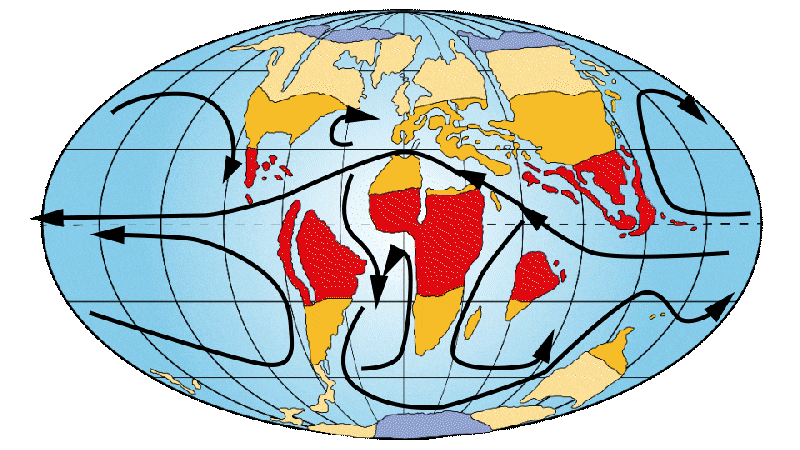
Figure 2: Distribution of continents, ocean currents and environments on Earth 65 Ma ago.
Between 56 and 34 million years ago
The continents move slightly. India moves closer to Asia and Australia takes off from Antarctica. This changes the way ocean currents are distributed around the globe. It was at this time that the so-called “Lower Eocene climatic optimum” occurred, corresponding to the warmest period of this age. Tropical zones then rise to much higher latitudes. It was at this time, around 56 Ma, that the first primates appeared.
Figure 3: Distribution of continents, ocean currents and environments on Earth 50 Ma ago.
Around 40 million years ago
Drake Strait, the arm of the sea between South America and Antarctica, is beginning to open up. This opening brings 2 oceans into contact, increasing the formation of diatoms and resulting in atmospheric change. Diatoms are photosynthetic microorganisms. This means they capture carbon dioxide (CO2) and release oxygen (O2) into the atmosphere, thereby reducing the amount ofCO2 in the atmosphere. The greenhouse effect is then less powerful and temperatures fall. This marked the beginning of a period of climatic destabilization and cooling. This major climate change resulted in a mass extinction of the first primates, dubbed the “Great Divide”. Nevertheless, one group of primates survived, the simiiformes, better known as “monkeys”, to which we Homo sapiens belong. Tropical forests are now confined to the equator, while temperate forests are spreading. This leads to new distributions of flora and fauna.
Figure 4: Distribution of continents, ocean currents and environments on Earth 30 Ma ago.
From 23 million years ago
The layout of the continents began to resemble what we know today. A large marine current, known as the circumpolar current, is finally being set up around Antarctica. This causes the oceans to shift from a warm to a cold regime. This change in currents led to the appearance of the first polar caps at the north and south poles. A greater fragmentation and diversification of climatic zones is taking place, similar to what exists today.
Figure 5: Distribution of continents, ocean currents and environments on Earth 20-16 Ma ago.
Setting up the Rift Valley
One last highlight before we wrap up! Around 10 Ma, the Rift Valley began to take shape. This corresponds to an almost 6,000 km-long fissure running from the Red Sea to the south of the African continent. The Rift Valley is thought to have played an important role in the emergence of the first Hominins, but we’ll tell you more about that another time!
We stop at the gateway to prehistory. This article is far from exhaustive on the Cenozoic, but our aim is to help you understand that the history of the Earth and life are intimately linked. Climatic, atmospheric and geological upheavals have a major influence on the distribution of plant and animal species, as well as on their extinction and appearance. So, like any other animal species, the emergence of hominins depends on the environment.
Feel free to ask us questions and give us feedback on the blog. You can also contact us by e-mail. You can also follow us on Instagram, Facebook, Twitter, LinkedIn, Youtube and TikTok!
See you soon,
The Prehistory Travel team.
Bibliography :
[1] Costeur L., Maridet O., Merceron G. (2018) -Les mammifères cénozoïques: Diversifications, adaptations et environnements, ISTE Group, 287 p.
[2] Friis E.M., Crane P.R., Pedersen K.R. (2011) – Early Flowers and Angiosperm Evolution, Cambridge University Press, 597 p.
[3] Huyghe D. (s. d.) – Changements climatiques globaux et forçage tectonique au Paléogène. Exemples du Bassin de Paris et des Pyrénées, , p. 359.
[4] Lagabrielle yves, Godderis yves, Donnadieu yannick, Malavieille J., Suarez M. (2009) – The tectonic history of Drake Passage and its possible impacts on global climate, Earth and Planetary Science Letters, 279, 3-4, p. 197-211.
Our solar system was born around 4.6 billion years ago (later abbreviated to Ga). It was formed within a large cloud of gas and dust. Under the effect of gravitation, the matter in this primitive cloud moved closer together, forming a ball of gas at its center. The temperature rose and a star, the Sun, began to shine. The rest of the primitive cloud remained orbitng the Sun. Bodies of matter gradually coalesced, giving rise to the planets a few million years later. The Earth was formed around 4.56 Ga ago.
How does life appear?
Nevertheless, the conditions necessary for the appearance of life on Earth were far from being met. Temperatures exceeded 100°C and our planet is covered by an ocean of magma. It took several million years before the temperature dropped and oceans and continents formed (between 4.5 and 4.4 Ga). The first life forms began to appear between 4 and 3.8 Ga. Around 4 Ga, organic matter formed from gases such as nitrogen, methane and carbon dioxide. This gives rise to proteins and nucleic acids, which are essential constituents of living organisms.
The first living organisms.
The oldest fossil traces of living organisms date back to 3.8 Ga with the stromatolites. These are sedimentary constructions resulting from the biological activity of very ancient bacteria. These bacteria photosynthesize by capturing carbon dioxide from the atmosphere and releasing oxygen. This type of bacteria still exists today. These are the cyanobacteria. The oldest traces of stromatolites have been found in Australia and Argentina. The emergence of life as we know it today is intimately linked to oxygen, a gas that is indispensable to many of today’s living organisms. Although it began to be released around 3.8 Ga, it wasn’t until around 2.6 Ga that the concentration of oxygen in the atmosphere became significant.
Conditions for the emergence of life
Three main conditions are necessary for the emergence of life as we know it: a continental crust, an oxic atmosphere and the presence of water in liquid form. Let’s not forget that one of our planet’s main characteristics is the abundant presence of liquid water. Indeed, for a long time to come, life continued to be confined to the aquatic realm. It was not until 400 million years ago (later abbreviated to Ma) that the first land animals emerged!
The mechanisms behind the appearance of the first organisms are unknown. Nevertheless, we do know that all living things today have a common origin, given that all living beings possess the molecules we mentioned earlier (proteins and nucleic acids, and also RNA and DNA). Scientists are obviously looking for the last common ancestor of all living things. This hypothetical ancestor is nicknamed LUCA, an acronym for Last Universal Common Ancestor. But be careful! LUCA is only the ancestor of today’s organisms. Other forms of life may have existed, but they simply didn’t leave any descendants.
Milestones in the history of life
Here are a few milestones in the history of life.
Figure 1: Timeline showing some of the major milestones in the history of life on Earth.
And what about human beings?
Human history began between 8 and 5 Ma, well after the appearance of our planet and the emergence of life on Earth! The time scale is dizzying, and this allows us to emphasize an important point: the notion of long and short time periods. 8 Ma may seem like a long time to us, but on the scale of our planet, it’s nothing at all! And it’s even shorter when we consider the emergence of the Homo genus 2.8 Ma ago or Homo sapiens around 300,000 years ago! Time is relative!
Don’t hesitate to ask us questions and give us your feedback on the blog. You can also contact us by e-mail. You can also follow us on Instagram, Facebook, Twitter, TikTok, Linkedin and YouTube!
See you soon,
The Prehistory Travel team.
Bibliography:
[1] Forterre P, Gribaldo S., “The origin of modern terrestrial life”, HFSP J., 2007, 1(3):156-68. doi: 10.2976/1.2759103.
[2] Forterre P., Gribaldo S., Brochier C. (2005) – ” Luca : à la recherche du plus proche ancêtre commun universel “, Med Sci (Paris), vol. 21, n°10, 2005.
[3] Grimaud-Hervé D., et al, History of ancestors. The great adventure of prehistoryEditions Errance, 2015
Pour offrir les meilleures expériences, nous utilisons des technologies telles que les cookies pour stocker et/ou accéder aux informations des appareils. Le fait de consentir à ces technologies nous permettra de traiter des données telles que le comportement de navigation ou les ID uniques sur ce site. Le fait de ne pas consentir ou de retirer son consentement peut avoir un effet négatif sur certaines caractéristiques et fonctions.
Fonctionnel
Always active
Le stockage ou l’accès technique est strictement nécessaire dans la finalité d’intérêt légitime de permettre l’utilisation d’un service spécifique explicitement demandé par l’abonné ou l’utilisateur, ou dans le seul but d’effectuer la transmission d’une communication sur un réseau de communications électroniques.
Préférences
Le stockage ou l’accès technique est nécessaire dans la finalité d’intérêt légitime de stocker des préférences qui ne sont pas demandées par l’abonné ou l’utilisateur.
Statistiques
Le stockage ou l’accès technique qui est utilisé exclusivement à des fins statistiques.Le stockage ou l’accès technique qui est utilisé exclusivement dans des finalités statistiques anonymes. En l’absence d’une assignation à comparaître, d’une conformité volontaire de la part de votre fournisseur d’accès à internet ou d’enregistrements supplémentaires provenant d’une tierce partie, les informations stockées ou extraites à cette seule fin ne peuvent généralement pas être utilisées pour vous identifier.
Marketing
Le stockage ou l’accès technique est nécessaire pour créer des profils d’utilisateurs afin d’envoyer des publicités, ou pour suivre l’utilisateur sur un site web ou sur plusieurs sites web ayant des finalités marketing similaires.